 Մեր հետազոտությունը կենտրոնանում է այն մոլեկուլային մեխանիզմների ընկալման վրա, որոնցով ռոդոպսինի մուտացիաները և դրա թրաֆիքինգը կարգավորող գեները հանգեցնում են ցանցաթաղանթի հիվանդությունների: Ռոդոպսինը ցանցաթաղանթային գավազանով ֆոտոընկալիչ բջիջների լույսի զգայուն թաղանթների հիմնական բաղադրիչն է, որտեղ այն գործում է որպես լույսի զգացող GPCR, որը նախաձեռնում է ֆոտոփոխանցման կասկադ: Ռոդոպսինը նաև հանդիսանում է ֆոտոռեցեպտորների առողջության կենտրոնական կարգավորիչ և ենթակա է ավելի քան 150 տարբեր մուտացիաների, որոնք առաջացնում են պիգմենտոզային ռետինիտ: Ռոդոպսին C- տերմինի վրա ազդող մուտացիաները առաջացնում են պիգմենտոզային autosomal dominant retinitis pigmentosa (ADRP) ծանր ձևեր: Մեր հետազոտության ընթացքում մենք սահմանեցինք ռոդոպսինի C- տերմինալ VxPx մոտիվը որպես պահպանված թարթիչների թիրախավորման պահպանված ազդանշան (CTS): Մենք հայտնաբերեցինք թարթիչավոր թիրախավորման համալիրի այն բաղադրիչները, որոնք ճանաչում են այս ազդանշանը `կարգավորելու ռոդոպսինների թրաֆիքինգը և ցանցաթաղանթային գավազանով ֆոտոռեցեպտորների թաղանթի նորացումը:
Մեր հետազոտությունը կենտրոնանում է այն մոլեկուլային մեխանիզմների ընկալման վրա, որոնցով ռոդոպսինի մուտացիաները և դրա թրաֆիքինգը կարգավորող գեները հանգեցնում են ցանցաթաղանթի հիվանդությունների: Ռոդոպսինը ցանցաթաղանթային գավազանով ֆոտոընկալիչ բջիջների լույսի զգայուն թաղանթների հիմնական բաղադրիչն է, որտեղ այն գործում է որպես լույսի զգացող GPCR, որը նախաձեռնում է ֆոտոփոխանցման կասկադ: Ռոդոպսինը նաև հանդիսանում է ֆոտոռեցեպտորների առողջության կենտրոնական կարգավորիչ և ենթակա է ավելի քան 150 տարբեր մուտացիաների, որոնք առաջացնում են պիգմենտոզային ռետինիտ: Ռոդոպսին C- տերմինի վրա ազդող մուտացիաները առաջացնում են պիգմենտոզային autosomal dominant retinitis pigmentosa (ADRP) ծանր ձևեր: Մեր հետազոտության ընթացքում մենք սահմանեցինք ռոդոպսինի C- տերմինալ VxPx մոտիվը որպես պահպանված թարթիչների թիրախավորման պահպանված ազդանշան (CTS): Մենք հայտնաբերեցինք թարթիչավոր թիրախավորման համալիրի այն բաղադրիչները, որոնք ճանաչում են այս ազդանշանը `կարգավորելու ռոդոպսինների թրաֆիքինգը և ցանցաթաղանթային գավազանով ֆոտոռեցեպտորների թաղանթի նորացումը:
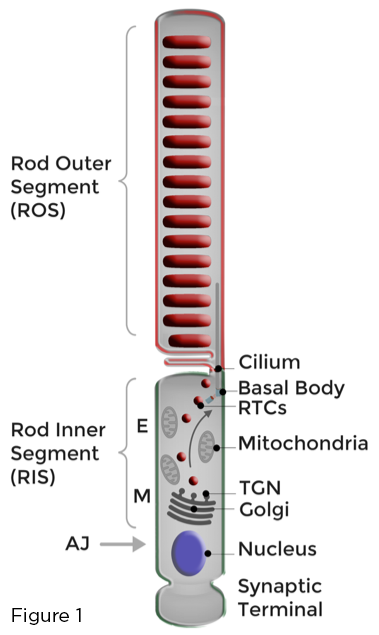 Ձողերի ֆոտոռեցեպտորների օպտիմալ գործառույթի հիմքը ռոդոպսինով հագեցած լուսազգայուն թաղանթների խիստ բաժանումն է գավազանի արտաքին հատվածը (ROS) ձևավորող յուրահատուկ ձևափոխված առաջնային թարթիչին: Ձողերի ֆոտոռեցեպտորների հոմեոստազը պահպանվում է թեթև վնասված ROS մեմբրանների շարունակական համալրմամբ և ֆոտոփոխանցման մեջ ներգրավված ROS սպիտակուցների և լիպիդների արդյունավետ սեկվեստրացիայի միջոցով, հեռու բջջի ներքին հատված (RIS) անունով հարակից բջջային մարմնում իրականացվող բազմազան բջջային գործառույթներից: կամ գավազանով սինապտիկ տերմինալում: Գոլջին և կենսասինթետիկ այլ օրգանոիդները տեղայնացված են ԱՊՀ -ում, միոիդային շրջանում (Մ): Բևեռացված թաղանթների թրաֆիքինգը ցանցաթաղանթային գավազաններում ներառում է ԳԻՆԳ-ի միջոցով ռոդոպսինի փոխադրիչների (ԳԹԿ) հսկայական քանակի սինթեզ, տեսակավորում և փոխադրում, ըստ ԱՊՀ-ի: RTC- ներն անցնում են էլիպսոիդային շրջանը (E), որը լցված է միտոքոնդրիայով և միաձուլվում են RIS պլազմային թաղանթով ՝ թարթիչի հարևանությամբ: Նոր սինթեզված թաղանթները այնուհետ հանձնվում են ROS- ին (Նկար 1):
Ձողերի ֆոտոռեցեպտորների օպտիմալ գործառույթի հիմքը ռոդոպսինով հագեցած լուսազգայուն թաղանթների խիստ բաժանումն է գավազանի արտաքին հատվածը (ROS) ձևավորող յուրահատուկ ձևափոխված առաջնային թարթիչին: Ձողերի ֆոտոռեցեպտորների հոմեոստազը պահպանվում է թեթև վնասված ROS մեմբրանների շարունակական համալրմամբ և ֆոտոփոխանցման մեջ ներգրավված ROS սպիտակուցների և լիպիդների արդյունավետ սեկվեստրացիայի միջոցով, հեռու բջջի ներքին հատված (RIS) անունով հարակից բջջային մարմնում իրականացվող բազմազան բջջային գործառույթներից: կամ գավազանով սինապտիկ տերմինալում: Գոլջին և կենսասինթետիկ այլ օրգանոիդները տեղայնացված են ԱՊՀ -ում, միոիդային շրջանում (Մ): Բևեռացված թաղանթների թրաֆիքինգը ցանցաթաղանթային գավազաններում ներառում է ԳԻՆԳ-ի միջոցով ռոդոպսինի փոխադրիչների (ԳԹԿ) հսկայական քանակի սինթեզ, տեսակավորում և փոխադրում, ըստ ԱՊՀ-ի: RTC- ներն անցնում են էլիպսոիդային շրջանը (E), որը լցված է միտոքոնդրիայով և միաձուլվում են RIS պլազմային թաղանթով ՝ թարթիչի հարևանությամբ: Նոր սինթեզված թաղանթները այնուհետ հանձնվում են ROS- ին (Նկար 1):
RTC- ների դասակարգումը կարգավորվում է Rab և Arf ընտանիքների փոքր GTPases կողմից, որոնք կենտրոնական դեր են խաղում ներբջջային թաղանթների թրաֆիքինգի կազմակերպման, ինչպես նաև թաղանթների առաջնային թարթիչներին հասցնելու գործում: Թարթիչների ձևավորման և պահպանման մեջ ներգրավված սպիտակուցները կոդավորված են ցանցաթաղանթային հիվանդության ժառանգական գեների մոտ 25% -ով, մուտացիաներն առաջացնում են ցանցաթաղանթի դեգեներացիա, կիստայական երիկամներ, գիրություն և նյարդային խողովակի արատներ գենետիկական լայն խանգարումների դեպքում, որոնք միասին կոչվում են կիլիոպաթիա:
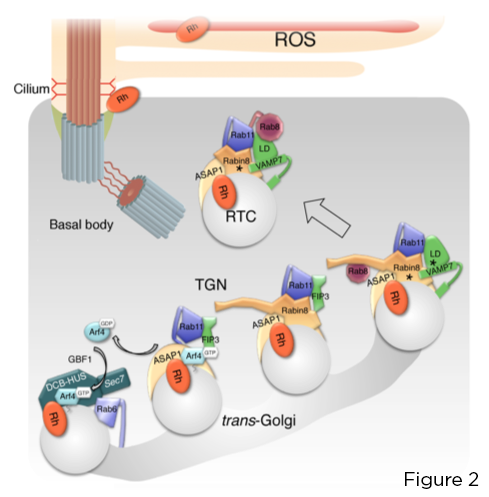 Փոքր GTPase Arf4- ը ճանաչում և ուղղակիորեն կապում է rhodopsin C- տերմինալը VxPx CTS- ին: Arf4- ը, որը ակտիվացված է Golgi ֆոտոընկալիչում Arf գուանին նուկլեոտիդների փոխանակման գործոնով (GEF) GBF1, նախաձեռնում է թիրախային կապի աստիճանական հավաքում, որը կենտրոնացած է Arf GTPase ակտիվացնող սպիտակուցի (GAP) ASAP1 վրա, որը միջնորդում է GTP հիդրոլիզը Arf4- ի և Rab11a- ի վրա: -FIP3-Rabin8 երկակի էֆեկտորային համալիր: Այս համալիրը վերահսկում է բարձր պահպանված Rab11a-Rabin8-Rab8 թիրախավորման մոդուլի հավաքումը, որն ուղղակիորեն հավաքագրում է R-SNARE VAMP7- ը RTC- ների վրա ՝ թարթիչային բազայում դրանց միաձուլումը կարգավորելու համար ՝ VAMP7- ի համակցված պլազմային թաղանթով SNAREs syntaxin 3 և SNAP-25 (Նկար 2):
Փոքր GTPase Arf4- ը ճանաչում և ուղղակիորեն կապում է rhodopsin C- տերմինալը VxPx CTS- ին: Arf4- ը, որը ակտիվացված է Golgi ֆոտոընկալիչում Arf գուանին նուկլեոտիդների փոխանակման գործոնով (GEF) GBF1, նախաձեռնում է թիրախային կապի աստիճանական հավաքում, որը կենտրոնացած է Arf GTPase ակտիվացնող սպիտակուցի (GAP) ASAP1 վրա, որը միջնորդում է GTP հիդրոլիզը Arf4- ի և Rab11a- ի վրա: -FIP3-Rabin8 երկակի էֆեկտորային համալիր: Այս համալիրը վերահսկում է բարձր պահպանված Rab11a-Rabin8-Rab8 թիրախավորման մոդուլի հավաքումը, որն ուղղակիորեն հավաքագրում է R-SNARE VAMP7- ը RTC- ների վրա ՝ թարթիչային բազայում դրանց միաձուլումը կարգավորելու համար ՝ VAMP7- ի համակցված պլազմային թաղանթով SNAREs syntaxin 3 և SNAP-25 (Նկար 2):
Rab11a-Rabin8-Rab8 թարթիչավոր կասկադի հիմքում ընկած է Rab8 GEF Rabin8- ը, բազմաֆունկցիոնալ փայտամածային սպիտակուցը, որը փոխազդում է ընտրված թարթիչավոր սպիտակուցների հետ, ինչպիսիք են TRAPPII թրաֆիքինգի համալիրը և BBSome- ը ՝ առաջարկելով կենտրոնական դեր զգայական ընկալիչների կիլյար ուղիներում: Նրա գործառույթի վրա ազդում են NDR2 kinase- ի (STK38L) մուտացիաները, որոնք ծածկագրված են ցանցաթաղանթի վաղ դեգեներացիայի (erd) գենով, որը համապատասխանում է մարդու ciliopathy Leber բնածին ամաուրոզին (LCA): Մեր ներկայիս ուսումնասիրությունը ցույց է տալիս, որ մարդկային GFP-Rabin8 արտահայտված տրանսգենիկ X. laevis համախմբվում է էնդոգեն Rabin8- ի և ռոդոպսինի հետ Golgi- ում և RTC- ներում ՝ ճանապարհ հարթելով ապագա ուսումնասիրությունների համար, որոնք ուղղված են Rabin8- ի դերին մեմբրանի առաջընթացում թարթիչային ուղու երկայնքով, որը պոտենցիալ խախտված է ցանցաթաղանթի ժառանգական դեգեներատիվ հիվանդությունների դեպքում:
Հավաքականորեն, մեր ուսումնասիրությունները պարզեցին, որ մեմբրանի թիրախավորումը դեպի ROS- ը թարթիչների թիրախավորման պահպանված ձև է: VxPx մոտիվը առկա է թարթիչային թաղանթի այլ սպիտակուցներում: Պահպանված Arf4- ի վրա հիմնված թիրախավորման համալիրը թիրախավորում է առաջնային թարթիչների զգայական ընկալիչները `փոքր GTPases- ի և դրանց կարգավորիչների բարդ ֆունկցիոնալ ցանցերի միջոցով, որոնք չափազանց զգայուն են ցանցաթաղանթների դեգեներացիաների և կիլիոպաթիաների պատճառած մուտացիաների նկատմամբ:
Deretic D., Lorentzen E և Fresquez T. (2019): Arf4- ի վրա հիմնված թարթիչային թաղանթ-թիրախավորման համալիրի նրբություններն ու ելքերը: Փոքր GTPases. Փոքր GTPases: 2019 մայիսի 9: 1-12: doi: 10.1080/21541248.2019.1616355: [Epub տպագրությունից առաջ]
Kandachar V, Tam BM, Moritz OL and Deretic D. (2018) Փոխազդեցության ցանց SNARE VAMP7- ի և Rab-GTPases- ի միջև ՝ թարթիչ թաղանթ-թիրախավորման համալիրի շրջանակներում: J Cell Sci- ն. (2018) J Cell Sci. 2018 թ. Դեկտեմբերի 10; 131 (24): pii: jcs222034. doi: 10.1242/jcs.222034.
Wang J., Fresquez T. Kandachar V. and Deretic D (2017): Arf GEF GBF1- ը և Arf4- ը սիներգիզացնում են զգայական ընկալիչ բեռը ՝ ռոդոպսինը, կարգավորելու թարթիչային թաղանթների թրաֆիքինգը: J Cell Sci- ն. J Cell Sci. 2017 թվականի դեկտեմբերի 1; 130 (23) ՝ 3975-3987: doi: 10.1242/jcs.205492. Epub 2017, հոկտեմբերի 12:
Vetter M, Wang J, Lorentzen E, and Deretic D. (2015) Rab11- էֆեկտորային փոխազդեցության ցանցի նոր տեղագրությունը թարթիչ թաղանթ-թիրախավորման համալիրի ներսում: Փոքր GTPases: 2015 թ. Հոկտեմբերի 2; 6 (4): 165-73: doi: 10.1080/21541248.2015.1091539: Epub 2015, սեպտեմբերի 23:
Wang J, and Deretic D. (2015) The Arf and Rab11 էֆեկտորը FIP3- ը գործում է սիներգիկորեն Arf GAP ASAP1- ի հետ ՝ Ռաբին 8 -ին ուղղորդելու թարթիչային ընկալիչների վրա: J Cell Sci- ն. J Cell Sci jcs.162925; Առցանց առցանց հոդված, 11 փետրվարի, 2015 թ., Doi: 10.1242/jcs.162925 (2015).
Wang J. and Deretic D (2014): Մոլեկուլային համալիրներ, որոնք ուղղորդում են ռոդոպսինի փոխադրումը առաջնային թարթիչներին: Prog Retin Eye Res. 2014 հունվար; 38: 1-19: doi: 10.1016/j.preteyeres.2013.08.004. Epub 2013, հոկտեմբերի 14
Wang J, Morita Y, Mazelova J and Deretic D. (2012): Arf GAP ASAP1- ը հարթակ է տրամադրում Arf4- և Rab11-Rab8 միջնորդավորված թարթիչավոր ընկալիչների թիրախավորումը կարգավորելու համար: EMBO J 31, 4057-4071:
Mazelova J, Ransom N, Astuto-Gribble L, Wilson MC and Deretic D. (2009) Syntaxin 3 և SNAP-25 զուգավորում, կարգավորվում է օմեգա -3 դոկոսահեքսաենաթթուով (DHA), վերահսկում է ռոդոպսինի առաքումը թարթիչների կենսածնության համար ստացված զգայական օրգանոիդներ, ձողի արտաքին հատվածներ: J. Cell Sci. 122, 2003-2013:
Mazelova J, Astuto-Gribble L, Inoue H, Tam BM, Schonteich E, Prekeris R, Moritz OL, Randazzo PA and Deretic D. (2009) Ciliary targeting motif VxPx ղեկավարում է թրաֆիքինգի մոդուլի հավաքումը Arf4- ի միջոցով: EMBO J. 28, 183-192:
Deretic D, Williams AH, Ransom N, Morel V, Hargrave PA and Arendt A. (2005) Rhodopsin C-terminus, ցանցաթաղանթի հիվանդություն առաջացնող մուտացիաների վայրը, կարգավորում է թրաֆիքինգը ՝ կապվելով ARF4- ի հետ: Proc. Նատլ. Acad. Sci. ԱՄՆ. 102: 3301-3306.